




2月29日,我校生命科学学院杨仲南/朱骏教授团队于《Plant Biotechnology Journal》发表了题为“Temperature and light reverse the fertility of rice P/TGMS line ostms19 via ROS homeostasis” (DOI:10.1111/pbi.14322) 的研究论文,该研究发现了一个新的水稻光温敏不育的新位点ostms19,揭示了水稻光温敏育性转换的共性机制。

光温敏不育作为一个重要的农艺性状,在水稻两系法杂交制种中发挥关键的作用。尽管在水稻中发现了诸多的光温敏不育位点,但是温度和光照如何影响水稻育性的共性机制尚不清楚。课题组在前期工作中利用模式植物拟南芥光温敏不育系的研究,解释了低温下小孢子发育缓慢是植物育性恢复的共性机制(Zhu et al., 2020, Nat Plant 6:360-367; Zhang et al., 2020, Plant Physiol 184: 923-932; Shi et al., 2021, Mol Plant 14:2104-2114; Wang et al.,2021, JIPB 64: 717-730)。但水稻中发现的光温敏位点与拟南芥有很大差异,提示水稻中除了缓慢生长外,还存在其他光温敏不育的共性机制。课题组发明了水稻光温敏不育系的创制技术,获得了一系列中花11(ZH11)为背景的水稻光温敏不育系。在2022年 (Zhang et al., 2022, PBJ 20:2023-2035;ostms18) 和2023年 (Han et al., 2022, PBJ 21:1659-1670;ostms15)发表两个具有直接育种价值水稻光温敏不育系的基础上,本次发表的是ostms19(图1)。

图1.水稻线粒体定位的PPR蛋白OsTMS19的缺陷导致温敏不育
OsTMS19基因编码一个水稻花药绒毡层表达的pentatricopeptide repeat (PPR) 蛋白。光温敏不育系ostms19中该基因620位碱基从T突变为C,导致氨基酸从Val突变为Ala。野生型水稻小孢子在花药发育的第10-11期,进行两次有丝分裂并开始合成花粉内壁。这个时期小孢子活性氧(ROS)非常敏感。高温不育条件下ostms19植株在这一时期检测到强烈的ROS信号,而在低温育性恢复条件下ROS能被有效清除,表明ROS稳态在温度调控ostms19过程中起重要的作用(图2A)。利用彗星实验检测发现,ostms19花粉细胞核DNA在高温条件损伤显著高于低温条件(图2B)。花粉内壁由小孢子(配子体)自身基因组控制。高温下ostms19花粉内壁缺陷导致不育,低温条件下内壁正常形成从而恢复育性(图2C)。遗传分析表明OsTMS19在孢子体中发挥作用,推测孢子体组织如绒毡层产生的ROS扩散到药室中,影响小孢子基因组以及花粉内壁形成缺陷从而呈不育表型,低温条件下ROS能被有效清除从而恢复育性。因此,ROS稳态在ostms19的光温敏不育中发挥着重要作用。
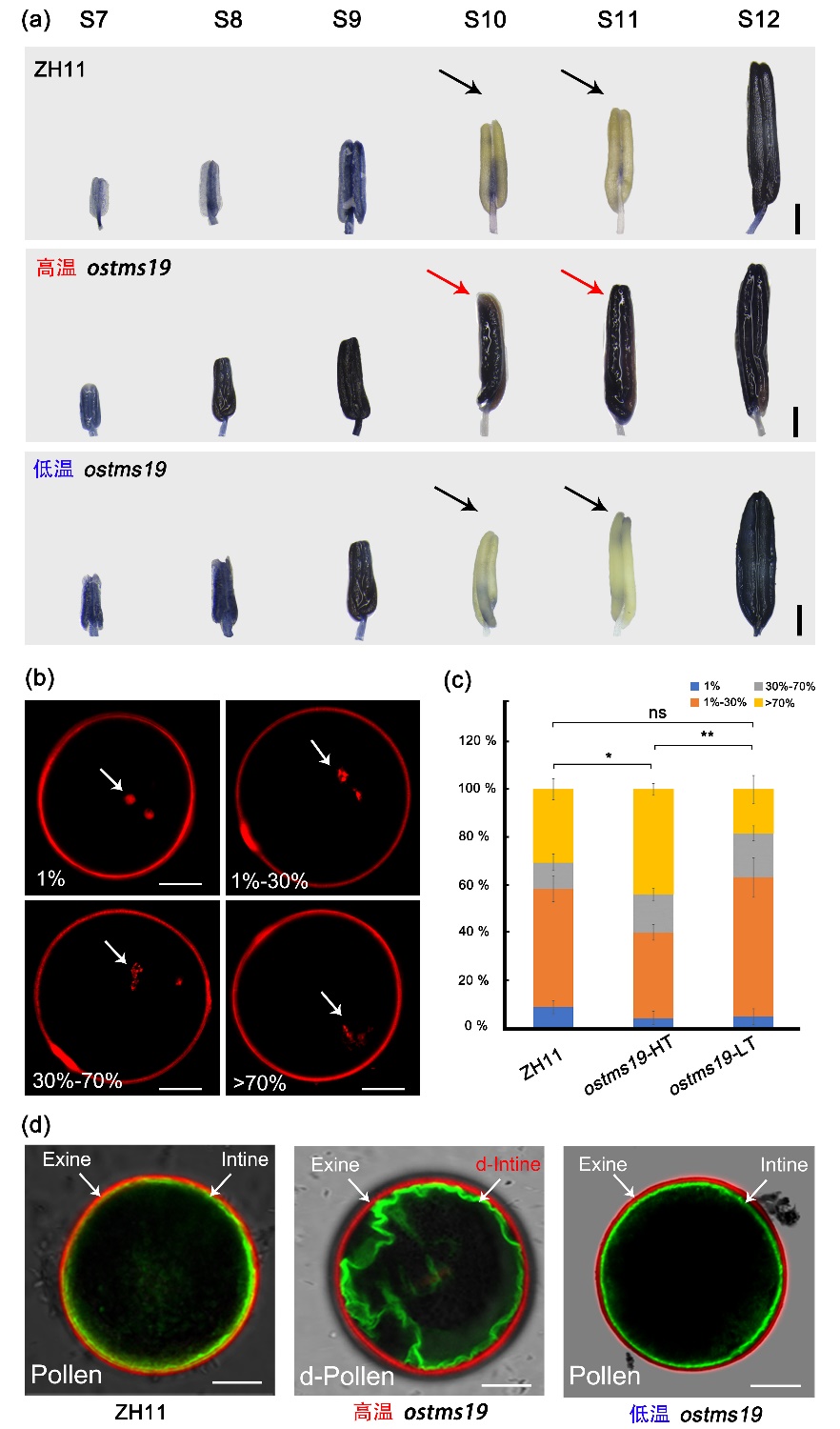
图2.温度通过ROS稳态调控ostms19花粉形成和育性
课题组随后分析了小孢子发育时期ROS稳态的改变是否也是其他水稻光温敏育性转换的共性机制。结果表明,在分析的所有三个水稻P/TGMS植株(tms5、ostms15和ostms18)中,高温下花药发育第10-11期的ROS也存在过量积累,低温条件下ROS能被有效清除(图3)。由此,课题组提出,响应温度和光照改变的ROS稳态变化可能是水稻光温敏不育系育性转换的共性机制。
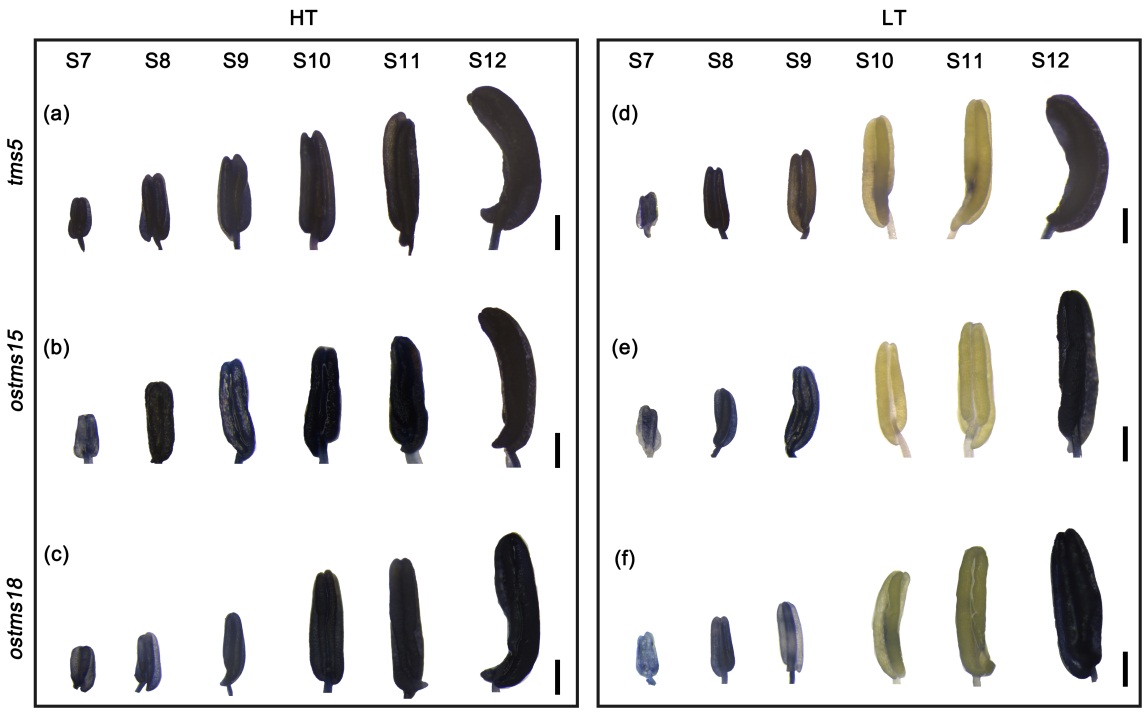
图3.不同P/TGMS品种花药发育10-11期都在高温积累ROS而在低温得到有效清除
结合先前模式植物拟南芥的研究成果,作者提出了作物光温敏育性转换的共性机制。1)花药发育花粉形成过程中经历一系列细胞壁的转换:四分体壁-花粉外壁-花粉内壁;2)光温敏基因直接或间接与这一细胞壁转换密切相关。光温敏不育系(P/TGMS)是相关基因突变引起;3)高温/长光照条件下的快速生长以及ROS过量积累,导致小孢子细胞壁无法有效转换而呈雄性不育;4)在低温/短光照下,缓慢发育降低了小孢子对细胞壁保护作用的要求,以及ROS的及时清除,保证了作物P/TGMS植株小孢子能发育成有功能的花粉从而恢复育性(图4)。
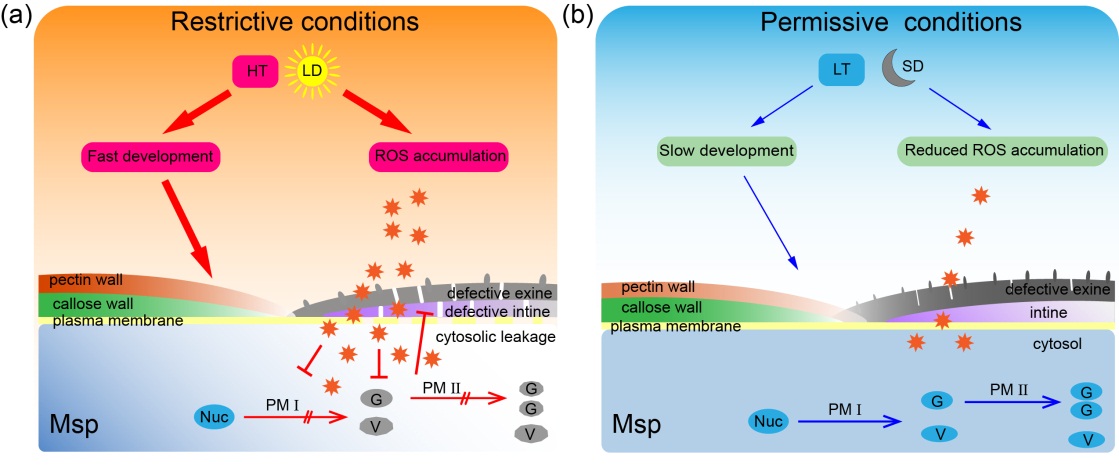
图4.水稻光温敏不育的恢复机制示意图
在该研究中,2024年新澳彩全年资料博士生周磊为第一作者,杨仲南教授与朱骏教授为共同通讯作者,黄学辉教授以及美国Donald Danforth Plant Science Center的Blake C. Meyers教授也参与了部分研究工作。该研究得到了国家重点研发计划,国家自然科学基金以及上海市教委和科委等课题的资助。
(供稿、摄影:生命科学学院)





 徐汇校区:上海市徐汇区桂林路100号
徐汇校区:上海市徐汇区桂林路100号